3.1. Effet du décocté de Zygophyllum album sur le glycogène hépatique
La quantité du glycogène hépatiques des rats à été déterminé sur l’organe prélevé et ce après le sacrifice des animaux. Les taux des glycogènes hépatiques des rats des 3 lots (témoins, diabétiques non traités et diabétiques traités par le décoté) sont représentés dans le tableau VII et illustrés dans la figure 17.
Tableau VII : Quantité du glycogène hépatiques (mg/100g de foie) des rats des 3 lots.
 des rats des 3 lots.")
")
Figure 17 : Taux du glycogène hépatique (mg/100g de foie) chez les rats des lots témoins, diabétiques témoins et diabétiques traités par le décocté de Z. album.
(Test de Student: * p <0.05, Diabétiques non traité vs témoins, *** p<0,001, diabétiques non traités vs diabétiques traités)
L’analyse de la figure, révèle que le taux du glycogène hépatique chez les rats du lot diabétiques non traités est diminué de 33 % et ce d’une manière significative (p < 0.05) comparé aux taux du glycogène hépatique des rats du lot témoins non diabétiques. Des résultats similaires ont été rapportés dans la littérature, en effet Soussi et al. (2009) ainsi que Suganya et al. (2012) ont observé que le glycogène hépatique des rats alloxanisés (diabétiques) a diminué respectivement de 50 % et de 70 % comparativement aux rats témoins. Ces résultats traduisent un moindre stockage du glycogène au niveau du foie des rats rendus diabétiques. L’injection d’alloxane engendre une réduction de l’insulinosécrétion qui a été observée par Kebièche et al. (2011). Cette insulinopénie pourrait être à l’origine de cette dégradation du glycogène, car l’insuline agit sur le foie en inhibant la glycogénolyse [Girard., 2008].
Par contre, chez les rats du lot diabétiques traités, le taux du glycogène hépatique est augmenté d’une façon très hautement significative (p < 0.001) par rapport au glycogène hépatique des rats du lot diabétiques non traités. Nous notons aussi que la quantité du glycogène hépatique des rats diabétiques traités est significativement (p < 0.05) supérieure à cellle des rats témoins. Cet effet a été déjà observé avec d’autres plantes considérées comme antidiabétiques. En effet Kebièche (2009) a rapporté une augmentation du glycogène hépatique sur des rats diabétiques traités par des flavonoïdes extraient d’une plante Ranunculus repens L ; il en est de même pour Soussi et al. (2009) avec Eucalyptus globulus.
Ainsi, à travers ces résultats, nous suggérons que le décocté de Zygophyllum album favorise le stockage du glycogène au niveau du foie. Selon El Ghoul et al. (2011), cette plante agirait en augmentant l’utilisation du glucose par certains organes cibles de l’insuline notamment le foie et de ce fait favoriserai la glycogénogenèse hépatique.
3.2. Effet du décoté de Zygophyllum album sur les lipides hépatiques
3.2.1. dosage des lipides totaux au niveau du foie
Les résultats du dosage des lipides totaux hépatiques des rats des 3 lots sont illustrés dans le tableau VIII et la figure 18.
Tableau VIII : Quantité des lipides totaux hépatiques (mg/100g de foie) des rats des 3 lots.
")
")
Figure 18 : Taux des lipides totaux hépatiques (mg/100g de foie) chez les rats des lots témoins, diabétiques non traités et diabétiques traités par le décocté de Z. album.
(Test de Student: ** p <0.01, Diabétiques non traité vs témoin ; ns, diabétiques non traités vs diabétiques traités)
A travers ces résultats, nous relevons que le taux des lipides totaux hépatiques des rats du lot diabétiques non traités est augmenté de 27 % et ce de manière hautement significative (p < 0.01) comparé au rat du lot témoins non diabétiques et cela nous fait suggéré que ce diabète expérimental (alloxanique) a induit une accumulation accrue des lipides au niveau du foie des rats diabétiques. Ces résultats rejoignent ceux de Dhandapani et al. (2002) et Prince et al. (2004) qui ont trouvés des taux de lipides au niveau du foie des rats diabétiques significativement élevés par rapport aux rats témoins non diabétiques.
Chez les rats diabétiques traités par le décocté de Z.album, nous constatons que les lipides totaux hépatiques sont diminués de 24.5 % par rapport aux rats du lot diabétiques non traités (p > 0.05). Donc le traitement par le décocté de Z. album à permis une réduction de l’accumulation des lipides totaux dans le foie des rats diabétiques traités. Ceci peut être la conséquence directe de la diminution du glucose sanguin (utilisation par les tissus périphériques) engendrée le Z.album limitant ainsi l’apport de substrat nécessaire (acetyl CoA) à la lipogenèse au niveau du foie ainsi qu’a l’inhibition de la lipolyse par l’insuline dont la sécrétion est potentialisé par le Z. album et donc privant le foie d’un excès d’acides gras libres.
3.2.2. Dosage des triglycérides hépatiques
Les résultats obtenus de la quantification des triglycérides au niveau du foie sont représentés dans le tableau IX et illustrés dans la figure 19.
Tableau IX : Quantité des triglycérides hépatiques (mg/100g de foie) des rats des 3 lots.
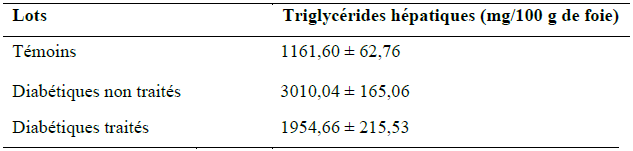
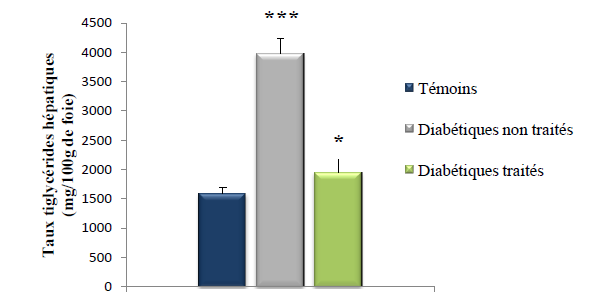
Figure 19 : Taux des triglycérides hépatiques (mg/100g de foie) chez les rats des 3 lots.
(Test de Student: *** p <0.001, Diabétiques non traité vs témoin ; * p < 0.05, diabétiques non traités vs diabétiques traités)
Ces résultats montrent que la quantité des triglycérides au niveau du foie des rats du lot diabétiques non traités est augmentée de 62 % et ce de manière très hautement significative (p < 0.001) comparée aux rats du lot témoins qui sont non diabétiques. Ces données sont en accord avec les travaux de Dhandapani et al. (2002). Cette synthèse accrue des triglycérides au niveau du foie pourrait être expliqué par la réduction de la sécrétion d’insuline par les cellules β suite à l’injection d’alloxane aux rats du lot diabétiques non traités rapportés par Kebièche et al. (2011) ; Saha et al. (2012), ce qui engendrera une moindre inhibition de la lipolyse au niveau du tissu adipeux et une augmentation des acides gras libres plasmatiques qui afflueront vers le foie pour une synthèse des VLDL-triglycérides. cette synthèse se trouve non freinée suite à la réduction de l’insulinémie, car l’insuline exerce un effet inhibiteur sur la production hépatique de triglycérides par diminution du taux de traduction de l’ARN messager de l’ apo-B qui est la protéine de transport des VLDL et par inhibition de la lipase hormonosensible au niveau du tissu adipeux limitant ainsi les substrats nécessaire (AGL) à la formation des triglycérides au niveau du foie [Vergès,. 2007]
En revanche, chez les rats du lot diabétiques traité par le décocté du Z. album, nous notons que les triglycérides hépatiques sont diminués de 50 % (p < 0.05) par rapport aux triglycérides hépatiques des rats du lot diabétiques non traités. Ce résultat prouve que le traitement a fait décroitre la synthèse des triglycérides dans le foie des diabétiques traités.
L’un des mécanismes pouvant expliqué la différence au niveau de la synthèse hépatique des triglycérides entre les rats diabétiques et les rats diabétiques traités est celui avancé par El Ghoul et al,. (2011) qui supposent que le Z. album augmenterait la sécrétion de l’insuline qui inhiberai la lipolyse au niveau du tissu adipeux et freinerai la production hépatiques des triglycérides.
Effet du décocté de Zygophyllum album sur la peroxydation lipidique au niveau des organes
La peroxydation des lipides au niveau des organes est un des dommages causé par le stress oxydant. Cette peroxydation à été étudié par la quantification du malondialdéhyde (MDA) au niveau du pancréas et du foie.
3.2.3. Quantification du MDA pancréatique
Les taux du MDA au niveau du pancréas des rats des 3 lots (témoins, diabétiques témoins et diabétiques traité par le décoté du Z. album) sont représentés dans le tableau X et illustrés dans la figure 20.
Tableau X : Quantité du MDA pancréatiques (μmole/ 100g de pancréas) des rats des 3 lots.
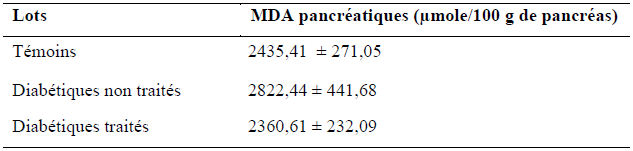
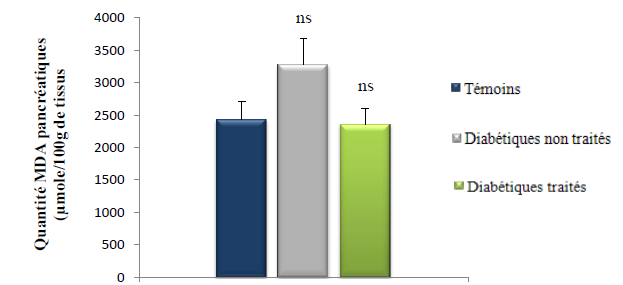
Figure 20 : Quantités du MDA pancréatiques (μmole/100g de foie) des lots : témoins, diabétiques non traités et diabétiques traités.
(Test de Student: ns p > 0.05, Diabétiques non traité vs témoin ; ns p > 0.05, diabétiques non traités vs diabétiques traités)
La lecture de la figure montre que le taux du MDA pancréatiques chez les rats diabétiques non traités est augmenté de 25 % par rapport aux rats du lot témoins (p > 0.05).
Cette peroxydation lipidique au niveau du pancréas pourrait être le résultat de à l’action l’alloxane qui est un générateur de radicaux libres [Lenzen,. 2008] ainsi qu’à l’hyperglycémie résultante du diabète expérimental. Ce stress oxydant pancréatique s’avère moins important comparé à d’autres travaux issus de la littérature, en effet Kebièche et al,. (2011) ; Chaudhry et al,. (2007) ont trouvé respectivement des taux de MDA pancréatique chez les rats alloxanisés 75 % et 46 % supérieurs à celui des témoins (p < 0.05), cela pourrait être expliqué par une hyperglycémie moins importante chez les rats de notre lot diabétiques non traités ce qui à contribué à une moindre aggravation du stress oxydant au niveau de leurs pancréas.
Par ailleurs, le traitement des rats du lot diabétiques traités par le décocté de Z. album à permis la réduction du taux du MDA pancréatique de 28 % comparativement aux rats du lot diabétiques non traités (p > 0.05). Nous remarquons aussi que le taux du MDA pancréatique des rats diabétiques traités est plus ou moins équivalent aux taux du MDA pancréatique des rats témoins. Ce qui prouve que le décocté de Z. album a limité la peroxydation lipidique à un niveau relativement normal.
Ces données rejoignent partiellement les résultats obtenus par El Ghoul et al. (2011) qui ont trouvés que le décocté de Z. album réduisait la peroxydation lipidique au niveau du pancréas des rats diabétiques de 35 % mais d’une façon significative (p < 0.05). Cette effet antioxydant pourrait être expliqué de deux manières : par la présence de flavonoïdes antioxydants dans les parties aériennes de Z. album [Sameh et al., 2011] et par la capacité de Z. album à augmenter les concentrations sériques des enzymes antioxydantes : superoxyde dismutase, glutathion peroxydase et catalase ainsi que des vitamines : E et C [El Ghoul et al., 2011].
3.2.4. Quantification du MDA hépatique
Les taux du MDA au niveau du foie des rats des 3 lots (témoins, diabétiques témoins et diabétiques traité par le décoté du Z. album) sont représentés dans le tableau et la figure.
Tableau XI : Quantité du MDA hépatique (μmole/ 100g de foie) des rats des 3 lots.

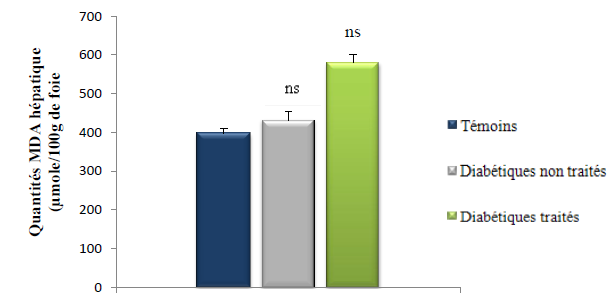
Figure 21 : Quantités du MDA hépatique (μmole/100g de foie) des rats des 3 lots.
(Test de Student: ns p > 0.05, Diabétiques non traité vs témoin ; ns p > 0.05, diabétiques non traités vs diabétiques traités)
D’après les résultats illustrés dans la figure, le taux de MDA au niveau du foie des rats diabétiques non traités est élevé modérément de 8 % comparés aux rats du lot témoins ; cette différence est non significative (p > 0.05). Les résultats de travaux d’autres auteurs : Kebièche et al. (2011), El Ghoul et al. (2011) ; Chaudhry et al. (2007) sont en désaccord avec nos résultat car ils ont mis en évidence une peroxydation lipidique significative au niveau du foie (p < 0.05) des rats alloxanisés comparés aux rats témoins. La faible peroxydation lipidique au niveau du pancréas de nos rats diabétiques non traités peut être le résultat d’un diabète moins important qui n’a pas aggravé d’une manière significative le stress oxydant hépatique.
On ce qui concerne les animaux du lot diabétiques traités, le MDA hépatiques est augmentés de manière non significative (p > 0.05) par rapport aux MDA hépatiques des rats du lot diabétiques non traités et du lot témoins. Ces donnés démontrent que le décocté de Z. album n’a pas permis la réduction de la peroxydation lipidique au niveau du foie de nos rats diabétiques traités et ces résultats sont en désaccord avec ceux de El Ghoul. J et al, (2011) qui ont conclus que le décocté de Z.album réduisait de manière significative (p < 0.05) le stress oxydant au niveau du foie des rats diabétiques traités. Nos résultats contradictoires (taux de MDA hépatique élevé chez les rats diabétiques traités par le Z. album) peuvent être liés à soit à une dose administrée de décocté qui été insuffisante ou à des artefacts formés au cours des manipulations et qui ont réagit avec le TBA.