Un stock d’une espèce donnée est dynamique dont les gains (entrées) sont représentés par le recrutement annuel et la croissance alors que les pertes (sorties) sont dues à la mortalité totale (Z). Cette dernière offre à décrire une mortalité par pêche (F) engendrée par les différentes opérations de pêches et la mortalité naturelle (M) qui exprime la mortalité due à toute cause autre que la pêche (Bouaziz, 2007).
5.1. Déterminations des paramètres de l’exploitation
En biologie des pêches, la manière la plus utile pour exprimer la décroissance en nombre des poissons d’un groupe d’âge au cours du temps est d’utiliser des taux exponentiels de décroissance (Pauly, 1985).

5.1.1.Estimation de la mortalité totale (Z)
Z est défini comme le coefficient instantané de mortalité totale, avant d’estimer séparément la mortalité par pêche et la mortalité naturelle, il est commode d’évaluer la mortalité totale (Gulland, 1969).
Nous avons utilisé quatre méthodes pour estimer le coefficient de mortalité total (Z).
5.1.1.1. Méthode de Powell-Wetherall (1986)
Cette méthode, déjà décrite dans la partie qui concerne l’étude de la croissance linéaire, permet l’estimation de Z/K ; connaissant K, le coefficient instantané de mortalité totale Z est alors déterminé.
5.1.1.2. Méthode de Jones et Van Zalinge (1981, in Sparre et Venema, 1996)
Les logarithmes népériens des captures cumulées par le bas (Nicum) sont portés sur un graphique en fonction des logarithmes népériens de la différence (L∞ – Li). La distribution de fréquences de taille est utilisée avec un intervalle de classe constant.
L’équation s’écrit :
![]()
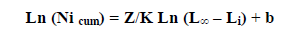
Dans cette expression Li représente la limite inférieure de la classe de rang i. Les paramètres de cette droite de pente Z/K, sont déduits du calcul de la régression linéaire.
5.1.1.3. Méthode de Beverton et Holt (1956)
")
5.1.1.4. Méthode de Pauly (1984)
Elle présente un double avantage : en plus de l’estimation de Z, elle permet l’évaluation de la taille de sélection.
L’auteur propose de représenter sur un graphique le logarithme népérien de la fréquence relative (%ni) en fonction de l’âge relatif (ti’).
Soit : Ln (%ni) = a ti’ + b
ni : l’effectif des individus pêchés dans la classe de taille de rang i (fréquence relative).
ti’ : l’âge relatif par classe de taille.
a et b : paramètres de la droite de régression.
L’âge relatif ti’ est calculé à partir de l’expression de Von Bertalanffy, avec la valeur de to = 0 (facultatif dans l’estimation de Z), soit
ti’ – to = – (1/K) [ Ln (1 – Li’/ L∞) ]
Li’ : le centre de classe de taille.
La courbe obtenue présente deux parties, seule la partie descendante est utilisée pour l’estimation de Z. Cette portion représente les classes d’âge entièrement capturables. La partie ascendante représente les classes d’âge incomplètement capturées, elle permet l’estimation de la taille de sélection (L50) (Hemida (2005, in Harchouche, 2006)).
Les calculs de ces trois dernières méthodes ont été réalisés par le logiciel (FISAT II).
5.1.2.Estimation de la mortalité naturelle (M)
Le coefficient instantané de mortalité naturelle M est l’un des paramètres les plus difficiles à évaluer. Ce coefficient exprime la mortalité due à toute cause autre que la pêche (prédation, maladies ou vieillissement).
5.1.2.1. Méthode de Pauly (1980 In Nouar, 1985)
La mortalité naturelle chez les poissons est corrélée avec la longévité, donc avec K. De même, elle est corrélée avec la taille puisque les grands poissons ont, en règle générale, moins de prédateurs que les petits. Nous pouvons également montrer que la mortalité naturelle chez les poissons est corrélée avec la température moyenne de leur environnement.
L’équation empirique de Pauly est donnée par la formule suivante :
Log10M = -0,0066-0,279 Log10L∞ + 0,6543 Log10K + 0,4634 Log10T
5.1.2.2. Méthode de Djabali et al. (1993)
L’équation de Djabali et al. (1993), déduite de régressions linéaires multiples, est basée sur les paramètres de croissance et de mortalité de 56 stocks de poissons vivants en Méditerranée (Harchouche, 2006).
Log10 M = – 0,0278 – 0,1172 Log10 L∞+ 0,5092 Log10 K
5.1.2.3. Méthode de Taylor (1959, In Harchouche, 2006)
En partant des paramètres de croissance de Von Bertalanffy K et to, cette méthode peut donner une estimation du coefficient de mortalité naturelle (M).
La formule utilisée est la suivante : M = 2.996 K / (2.996 + K to).
5.1.3. Estimation de la mortalité par pêche (F)
Connaissant M et Z, le taux de mortalité par pêche peut être déduit de la relation : Z=M+F
5.1.4. Estimation du taux d’exploitation (E)
Le taux d’exploitation (E) est défini par Pauly (1985) comme étant la quantité :
E = F / (F + M) = F / Z
Ce taux permet à lui seul d’estimer (grossièrement) si un stock est surexploité ou non, à partir de l’hypothèse que la valeur optimale de E (Eopt) est voisine de 0,5; cette hypothèse, E ≈ 0,5 = taux d’exploitation optimal, repose elle-même sur une autre hypothèse, à savoir que la prise équilibrée potentielle est optimisée quand F ≈ M (Gulland 1971, In Pauly,1985).
5.1.5. Recrutement et sélection
Les poissons de tailles ou d’âges divers ne subissent pas tous la même mortalité par pêche: les petits peuvent s’échapper à travers les mailles d’un filet, ou ne pas se trouver dans la principale zone exploitée. Il est utile de distinguer l’élément provenant uniquement du comportement (au sens le plus large) des animaux mêmes – recrutement – et celui qui est dû à la sélectivité de l’engin de pêche (Gulland,1969). Le recrutement est le processus par lequel un groupe d’âge arrive pour la première fois dans une pêcherie. La sélection est le phénomène d’entrée progressive du poisson dans les captures (Bougis, 1976, In Harchouche, 2006).
5.1.5.1. Estimation de la taille de recrutement :
Dans cette étude il est impossible de déterminer la taille de recrutement, ceci est en cause de l’absence des individus de petite taille.
5.1.5.2. Estimation de la taille de sélection :
La méthode de Pauly (1984 a et b) considère la partie gauche de la courbe de capture linéarisée. Cette partie, ascendante, représente les individus partiellement capturés par l’engin de pêche. Une façon simple d’estimer combien d’individus manquent à chaque âge consiste à extrapoler la droite à partir de laquelle le coefficient de mortalité totale Z est estimé.
L’effectif réel du stock CT est proportionnel au nombre de poissons capturés C et la probabilité P pour qu’un poisson d’âge t soit présent sur les lieux de pêche s’estime à partir de

Cette probabilité de capture P représentée en fonction de la taille, permet d’estimer la taille de première capture correspond à la probabilité P = 0,5. Le calcul des tailles de capture est réalisé par le logiciel FISAT II. Une fois la taille de première capture (Lc) déterminée, l’âge de première capture (tc) est calculé à partir de l’équation de Von Bertalanffy (Kennouche, 2008)
5.2. Estimation du niveau d’exploitation
On réserve souvent cette appellation à des modèles prévisionnels et plus ou moins explicatifs, pouvant conduire à des simulations. Un modèle n’a jamais la prétention d’épuiser la réalité. Ce n’est qu’une simplification, toujours abusive dans l’absolu. S’il n’épuise pas la réalité, le modèle doit en retenir l’essentiel, suffisamment réduit pour être manipulable (Laurec et Le Guen, 1981). Les modèles globaux ne tiennent pas compte des paramètres biologiques. En effet, ils ne font intervenir que les statistiques de pêche (prise de pêche et effort de pêche), ce qui dans notre cas ne peut être réalisé (les statistiques de pêche étant inexistantes pour les espèces étudiées).
Les modèles analytiques ou structuraux permettent de gérer un stock de poisson par une étude du rendement par recrue et une simulation des changements qui affectent ce stock.
De ce fait, nous avons appliqué :
· Le modèle de Beverton et Holt (1966).
· Le modèle rétrospectif : Analyse de cohorte de Jones (1984) fondé sur la longueur.
· Le modèle prédictif de Thompson et Bell (1934) fondé sur la longueur.
5.2.1.Le modèle de Beverton et Holt (1966) :
Le modèle de Beverton et Holt (1966) exprime le rendement relatif par recrue (Y/R)’, en permettant de déterminer le rapport entre rendement et l’effort de pêche pour différentes tailles de premières captures, il appartient à la catégorie des modèles fondés sur la longueur (Sparre et Vanema, 1996). L’expression de Y’/R est
")
Cette expression utilise trois paramètres : M/K, Lc/L∞ et E.
Le FISAT II a permis l’application du modèle de Beverton et Holt (1966), celui-ci est utilisé afin d’évaluer le rendement et la biomasse relatifs par recrue.
La biomasse relative par recrue (B’/R) est évaluée à partir de la relation suivante :
(B’/R)= (Y’/R)/F
D’autres paramètres sont fournis dans ce modèle comme le Emax, E0.1 et E0.5 exprimés graphiquement.
Emax : Exploitation avec rendement productif maximum.
E 0.1 : Taux d’exploitation pour une augmentation de Y’/R de l/10éme par rapport à E=0
E 0,5 : Valeur de E sous laquelle le stock a été réduit de 50% de sa biomasse inexploitée.
5.2.2.Analyse de cohorte de Jones (1984) fondée sur la longueur
L’analyse des populations virtuelles ou AVP est une méthode faisant appel aux populations capturées ; elle consiste à analyser ce que l’on peut observer, la capture ; de manière à estimer la population qui devrait se trouver en mer pour produire cette capture.
La méthode de Jones (1984) fait appel à des « pseudo-cohortes » caractérisées par des paramètres constants. L’analyse des cohortes largement utilisée dans le domaine d’évaluation des stocks d’animaux aquatiques est considérée comme l’une des méthodes les plus performantes en matière de modélisation analytique ; elle repose sur l’analyse de structures démographiques des captures (répartition par âges ou par tailles)en admettant que le recrutement et les mortalités par pêche aux différents âges et tailles restent constants, elle permet une estimation des taux instantanés de mortalité par pêche (F), ces derniers mesurent la pression réelle exercée sur le stock (Ghorbel et al., 1997 (In Harchouche, 2006)).
L’analyse des populations virtuelles ou APV permet de décrire l’état d’exploitation et de faire des évaluations fiables de la gestion des stocks.
Les formules pour le calcul de la production et la biomasse par l’analyse des cohortes sont représentées dans le tableau 1.
Tableau 1 : Formules utilisées pour le calcul de la production et la biomasse par l’analyse de cohorte (Jones, 1984).
")
Les calculs sont réalisés à partir de la dernière classe et poursuivis jusqu’à la première.
Selon Jones(1984) le taux d’exploitation de la dernière classe (F/Z terminal) est égal à 0.5 (nous pouvons aussi considérer F terminal = 0.5. Le nombre final de survivants sera alors :
N (L1) Final = C (L1 – L∞) / (F/Z) Final
Nous calculons alors le N (L1) de la classe précédente, etc(Sparre et Venema, 1996).
5.2.3. Modèle prédictif de Thompson et Bell (1934) fondé sur la longueur
Le modèle de Thompson et Bell (1934) sert à prédire les effets des modifications de l’effort de pêche sur les futures productions et biomasses à partir des données historiques d’une pêcherie. Ce dernier est fondé sur l’analyse de pseudo-cohortes de Jones (1984) permettant d’évaluer l’impact d’une variation de l’effort de pêche après détermination des coefficients de mortalité et donc fournissant aux responsables des ressources halieutiques les informations réelles sur l’état du stock (Sparre et Venema, 1996).
L’application de ce modèle nécessite l’empreinte des données d’entrées à une analyse de cohortes fondée sur la longueur qui sont:
– Les mortalités par pêche par classe de longueur F (Li – Li+1).
– Le nombre de poisson dans la plus petite classe N (Li).
– Le facteur de la mortalité naturelle par classe de longueur H (Li – Li+i).
– Les coefficients de la relation taille-poids (a et b).
Le groupe d’équations suivant résume sous une forme générale les formules correspondant au Modèle.
Tableau 2: Formules utilisées pour le modèle de Thompson et Bell (1934)
")
Pour les méthodes de Jones(1984) et Thompson et Bell (1934), les calculs et les graphes ont été programmés sur le logiciel Excel.
Page suivante : Chapitre III : Résultats, interprétations et discussions